Ladda ner presentationen
Presentation laddar. Vänta.

1
Förädlingssynpunkter Dag Lindgren Ekebo 07-03-20 Foto: Darius vid vårt seminarium Ekebo 2004

2
Ämnen att föredra. Antytt vilka grupper det sorterar under. Enligt uppdrag från Ola Rosvall: Antal pollen i polymix (grupp 3); Polymix breeding (grupp 3 + grupp 2); Breeding without breeding (grupp 3 + grupp 2); Korsningssystem SPM, DPM, Polycross (grupp 3); Obalanser (grupp 3 m fl.).
; Polymix breeding (grupp 3 + grupp 2); Breeding without breeding (grupp 3 + grupp 2); Korsningssystem SPM, DPM, Polycross (grupp 3); Obalanser (grupp 3 m fl.)..")
3
Antal pollen i en pollencocktail?

4
Conclusion: Somewhat below 10 seem sufficient…

5
”Grupp 3” säger 25 pollen Beaktar variationer i pollenkvalité mellan olika kloner; Gör det möjligt att variera pollen (inom ”population”) med bibehållen kompatibilitet; Ofta är färre pollen acceptabelt, men merkostnaden är troligen liten; ”Vi” har hittills utgått från att dominansvariationen är 25% av den additiva. Om ”vi” skulle höja till >75% kan det motivera översyn; Finns dokument ”polymix” på webben.

6
Urval framåt i PMX Idé Lambeth 2001; gör urval framåt i polymix familjerna; klara upp föräldrarna i en polymix i efterskott med markörer, Detta skulle kunna tänkas vara överlägset ”huvudspåret” och bör därför utvärderas, även om jag tror att ”många korsningar” är bättre om man förbereder många mödrar och fäder för korsning. Information om fadern i polymixen utnyttjas. Vid konventionell polycross-avkommeprövning av modern är fadern enbart ett störande brus.

7
PMX-breeding; förslag till design Indela en bpop i 5 avdelningar; Från en avdelning väljs 50 mödrar som ympas och från en annan 25 fäder som skördas på pollen till en pollenmix; Polycross, nytt fältförsök; Fastställ fäder till lovande träd i fältförsöket; Välj 50 till mödrar och 25 till fäder. Valet sker med en variant av Group Merit Selektion; Etc., cirkeln är sluten!

8
Dag’s övervägande om selektion framåt i polymixavkommor Biologiskt, praktiskt och tekniskt möjligt med befintlig teknik, men troligen inte förädlingsekonomiskt: Det saknas nog lämpliga objekt för att starta urvalet (för ointressanta och få fäder i de pollenblandningsavkommor som nått fältet); Gör systemstudier (POPSIM); Gör nu några fpop avdelningar med PMX-korsningar i verkligheten. Bra studier fordrar att verkliga data och material är bra! Skillnaden mot gårdagens polycrossar ligger i överväganden av pollenmixar och dess matchning till mödrar. Merkostnaden att framställa dessa material blir minimal, eftersom de ligger nära vad som är huvudspåret nu. Om Skogforsk skapar materialet nu, så kan göra man göra skarpa selektioner om 10-15 år. Finare optimering och effektiva algoritmer bör inte göras utan skarpa data. De bör därför inte utveckla de kommande 10 åren.

9
Breeding without Breeding (Concept coined by El-Kassaby 2006,)
")
10
Idea Let Nature do the recombination! No controlled crosses any more! Get pedigree by markers instead of crosses!

11
Can be applied at full scale directly following a pilot study; No risk; Probably brand new and fastly expanding research area!; Some of the designs I show are likely to work with little marker development, but may not be good breeding economy. Others (in particular where it is required to identify both parents), may need considerable marker development.
, may need considerable marker development..")
12
Breeding Without Breeding: Breeding values without test families No experimental trees or plants; Neglible measurement costs; Reward of investment immediate!

13
Breeding values without testfamilies Seed orchard Forest Parents with markers Find the goodies Estimate breeding value for ability to get good progeny

14
Breeding without Breeding Applikation – Enkel Bara två komponenter i fält som skogsbruket har ändå: Fröplantager och skog! Inga korsningar; Inga klonarkiv; Inga tester; Inga försöksstationer….

15
Simple Breeding without Breeding Cycle Seed orchard Forest Group merit selection

16
Breeding without Breeding Modification of mass selection - low input breeding Select plus trees in the forest; Put grafts in a seed orchard; Harvest seeds; Grow a forest; Select plus trees; The cycle is closed!

17
Modified Mass-selection Gain cannot be higer than curve at a given diversity Modified from Lindgren and Wei 1993 0 1 Combined index (maximizes gain) Note that phenotypic (=mass) selection is on the optimising curve, thus no way to get more gain without giving up diversity! Mass selection (easy) Between family (exhausts diversity) Within family (conserves diversity) Gain 0.5 Diversity Max Min
 Between family (exhausts diversity) Within family (conserves diversity) Gain 0.5 Diversity Max Min.")
18
But will Breeding without Breeding build up relatedness? We doubt to apply simple methods now because we do not know what it will do to relatedness; With genotyping by markers we know!!!; Markers offer more reliable control of coancestry than today!

19
Applikation – selektion i OP familjer Om pollenmolnet är delvis förädlat från ett begränsat antal fäder (avkommeprövning med fri avblomning i plantage) kan förädling bedrivas ungefär som vid urval framåt i polycross. Tänk efter om det finns någon fröplantage med dåligt testade kloner som kan lämpa sig för avkommeprövning med fri avblomning följt av urval framåt.

20
Application Norway spruce long term breeding

21
“Conventional” Norway spruce breeding (20) Clonal test by far the best Select at age 20 Crosses Cloning of 20 seedlings Field test with 20 ramets from each The breeding cycle is complete!
 Clonal test by far the best Select at age 20 Crosses Cloning of 20 seedlings Field test with 20 ramets from each The breeding cycle is complete!")
22
Breeding without Breeding Application Norway spruce clonal (20) Select at age 20 Cloning of 20 seedlings Field test with 20 ramets from each The breeding cycle is complete! Harvest seeds from good mothers Analyze seeds Selects seeds with good fathers

23
Advantages: No artificial crosses No clone archives or top-grafting Robust, optimal testing design, nothing lost if BwB is never applied! Norway spruce flowers irregular, but nothing has to be done until a good cone set observed Spruce flowers also rather well at age 20 in good clima provided intensive thinning some years before Measurements done the seed maturation year Clonal propagation takes some years, but 80% of the breeding cycle is used for the gain generating testing and only 20% for improductive wait! The breeding cycle is reduced maybe years (the time for top-grafting and wait for crosses).
..")
24
Old clone tests Could be used to harvest the gains from old clone tests which has not been utilised before (Hilleshög, Mellansvenska)…
…")
25
Rekommendation för BWB Det är ganska troligt specialfall av BWB kommer till praktisk användning inom tio år; Genomslag kan ske ganska snabbt; Understöd forskning och stimulera forskning på ”skarpa” material.

26
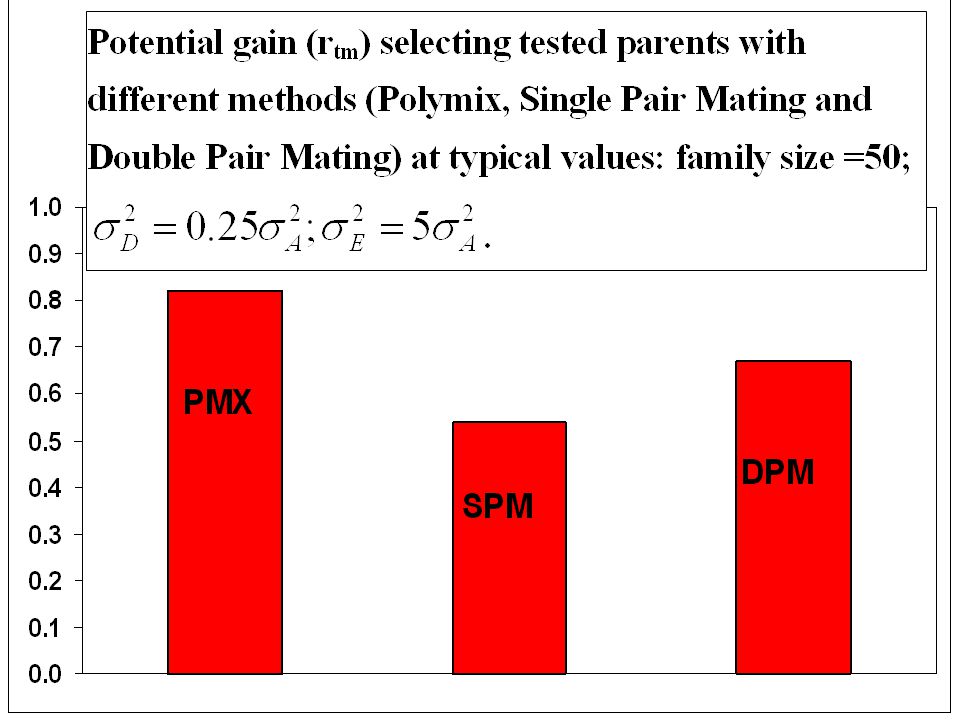
Korsningssystem DPM can remain as a general main design; SPM is not bad and can be used in low input situations or in lower strata of the breeding population; If one DPM fails and parents still get progeny in one cross it is not worth delaying crossing effort; SPM and DPM gives much information about BV of parents.

28
Selektion framåt i OP-familjer Om pollenmolnet är försök eller i fröplantager kan okänd fader trassla till, och kan man inte faderskapsbestämma med markörer så är jag tveksam; Ibland kan BWB tekniker vara användbara; Fortsatt betraktelse avser pollenmoln ur ”samma” population som modern valts, obesläktad med andra val.

30
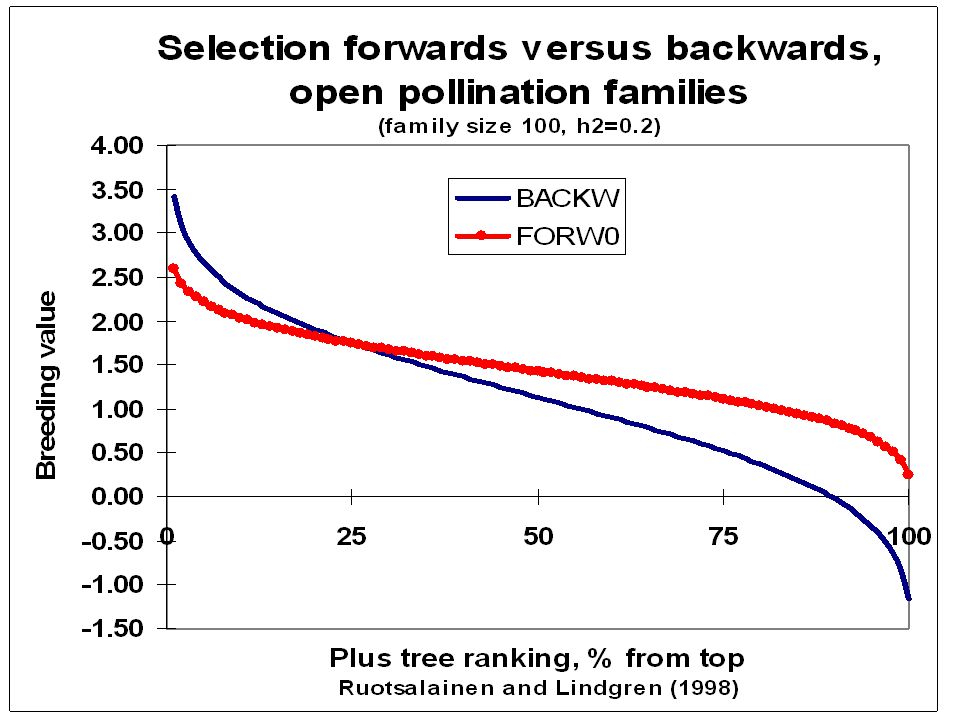
Forward selection in OP Parents ranking below 25 for BV in the BPOP can be replaced by the best OP offspring; For parents ranking higher than 25 continued efforts to get crosses is recommended; Anyway while waiting selections can be done from offspring; Several selections can be done from the OP offspring and OP progeny tested before crosses.

31
Slut grupp 2-3

32
Unbalances in Tree Breeding

33
“Genetic unbalances” are: The basis for evolution; Natural – balance is extremely unnatural; Unavoidable; Essence of breeding.

34
Genetic contributions varies. Natural selection favors some and disfavors others, (survival of the fittest), thus some contributions will increase, other decrease
, thus some contributions will increase, other decrease.")
35
Selected Not selected Selection means always unbalance, not selected components get no contribution Contribution of component I talk about more sophisticated unbalance

36
Svensk skogsträdsförädling 2007 kan ses som extrem obalans All avkomma till den andra generationen kommer från endast 20% av ”grundarna” de testade plusträden.

37
Different types of unbalances… Unbalance in genetic components (parents, founders); Unbalance in resources; Structure of breeding population (e.g. mating probability, PAM); Unbalances both at creating recruitment populations and at final selections, these have to be optimised together. Often they come together, thus “Nucleus” (elite-main) has many components of unbalance
; Unbalances both at creating recruitment populations and at final selections, these have to be optimised together. Often they come together, thus Nucleus (elite-main) has many components of unbalance.")
38
Why unbalance? Breeding has to consider: Gain; Gene diversity; Cost; Time; Interaction breeding → seed orchards. Unbalances may make the breeding system more optimal and efficient. Unbalances offer more degrees of freedom for optimization (balance is a form of simplification fundamentalism)
.")
39
Reasons against unbalance? Unbalance may just make things worse if not done wisely and skillfully; Unbalance is more demanding on management skills; Often the tools for handling unbalance are badly developed! Seldom transparent, difficult to sell; Sometimes the advantage is small, usually limited (3- 8%) and seldom drastic (20%); Advantages predicted for an ideal situation, usually somewhat less in the real world! Historically difficult to manage, thus traditional wisdom is against, now computers can do everything!!!?.... Requires calculations to be done.
 and seldom drastic (20%); Advantages predicted for an ideal situation, usually somewhat less in the real world. Historically difficult to manage, thus traditional wisdom is against, now computers can do everything!!! .... Requires calculations to be done..")
40
Unbalance is a black box! Requires lots of competence! Risky… Output: Gain, effective number, remaining ramets per clone Input: breeding values, ramet numbers

41
More reasons for unbalance Even a limited extra gain (e.g. 5% increase in gain) means enormous economic returns; Complete balance is practically unrealistic! Unbalance must anyway be managed, so why not do it efficient!?
 means enormous economic returns; Complete balance is practically unrealistic. Unbalance must anyway be managed, so why not do it efficient! .")
42
Quantitative evaluation of unbalance often overestimates the practical benefit! Genetic parameters (genetic correlations) change over time and environment; Environment changes; Unreliable parameter estimations; Breeding goals change and are not exactly predictable; Planned unbalances are influenced by unplanned;
 change over time and environment; Environment changes; Unreliable parameter estimations; Breeding goals change and are not exactly predictable; Planned unbalances are influenced by unplanned;.")
43
Suggestion: Apply unbalance, but with moderation! Apply unbalances; But do it with moderation and not too drastic; It might sometimes be a good idea to try compromising between balance and the predicted optimal unbalance; After gaining experience of unbalance, a larger share of predicted advantages may be utilized.

44
Unbalances in production population Simplest case, only unbalances in different contributions (e.g. clones, parents) matter. Jag gissar att detta nu är accepterat i Sverige så jag säger inte mer….
 matter. Jag gissar att detta nu är accepterat i Sverige så jag säger inte mer…..")
45
Breeding value of clone Contribution of clone A “Balance” More clones with different contributions can result in both more gain and more diversity! B C D E A “Unbalance” H G B D F C E Equal (balanced) contribution of clones!
 contribution of clones!.")
46
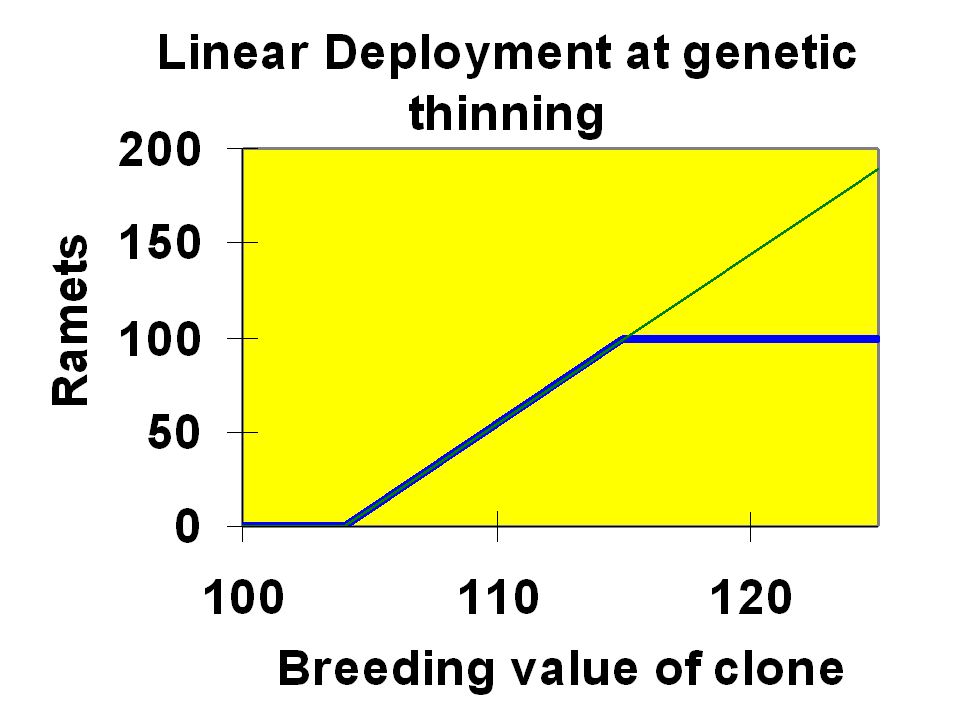
Linear deployment is optimal for establishment! Relate contribution linearly to breeding value; No other deployment combines higher gain with higher effective number.

48
At thinning ramets cannot be added, just withdrawn. Thus there is a highest number of ramets! Linear deployment works with constraints also

50
The “Swedish” model In the following, many of presented figures intend to be relevant for Sweden or the Swedish breeding strategy; Heading for a number of long term “breeding populations”, each of size 50; Heading for balance: Within family selection; Each parent get two full sib families; One selection per family. Start with tested plus trees (typical 200 per breeding population); Test recruitment population (clone-testing or progeny- testing); Genetic parameters, costs and time estimates should be relevant.
; Test recruitment population (clone-testing or progeny- testing); Genetic parameters, costs and time estimates should be relevant..")
51
Unbalanced contributions at the initiation of a tree improvement program Closing the breeding population is irreciprocal and can not be undone! Argument to play on the safe side!

52
Method developed for “optimal selection” in a population with family structure (can be visualized as unrelated full sibs with plus tree parents); Optimal selection among individuals with a family structure is close to linear deployment from parent offspring; Seems reasonable to start with crosses from about 150 plus trees to start up a breeding population; Differences from current Swedish program: no testing (“phenotypic selection”), thus heritability not high; no initial knowledge of plus tree breeding values. Generalizations from Wei PhD thesis (Wei 1995)
.")
53
Unbalances in setting up the first recruitment population Generalized from Ruotsalainen (2002) PhD thesis; An approximation to linear deployment (3,2,1).
 PhD thesis; An approximation to linear deployment (3,2,1).")
54
Unbalanced (60 founders)Balanced (50 founders) Rank of plus tree ProgeniesRank of plus tree Progenies 1-1031-502 11-302- 31-601- 61-200051-2000 Gain = selection intensity 1.3681.271 The same resources, the same gene diversity. But 8 percent more gain with unbalance!

55
Unbalance, eight percent more gain than balance! Share of represented founders (tested plus trees) Progenies per founder Best 1/63 Medium 1/32 Bottom 1/21
 Progenies per founder Best 1/63 Medium 1/32 Bottom 1/21.")
56
Result Andersson PhD thesis 1999 Unbalanced selection is superior to balanced in the initiation of a breeding program!

57
Unbalance by refreshing in F 1 Inspired from Andersson PhD thesis (1999) Unbalance could perhaps be introduced in F 1 by refreshing (adding formerly discarded plus-tree genes): In model-calculations it was favorable to replace 5-10% of founders at F 1 with new plus trees; That indicates that it may sometimes be beneficial to replace one or a few of bottom ranking BP members with new founders in the Swedish breeding; The introduced founders may have slightly lower BV, but the Group merit of the BP could increase. That would mean that a few F 1 BP would be crossed with new founders to form the next BP generation; The bottom ranking selected founders are only slightly superior to the best non-selected candidates. The refreshment may be e.g. old grafts in clonal archives with top BV ranks of those which have not F-1 progeny yet and which are in breeding populations there 50 or less parents has formed F1-progeny, which may serve as mothers for freshly collected pollen from F1-trees in the forest.

58
Unbalances in long term breeding

59
Wei (1995) demonstrated the potentially disastrous effects to use the strongly unbalanced selection resulting from maximizing breeding value in each generation; When gene diversity is exhausted, genetic gain drops; Sanchez (2000) studied the effect of a slight unbalance with quantitative simulation and small populations. It was noted that a slight unbalance often was more favorable in breeding than complete balance.

60
Results generalized from PhD thesis Rosvall (1999) Used POPSIM (tree improvement simulator) to study different aspects of long term breeding with simulation of a program similar to Swedish Norway spruce breeding. The capacity of the breeding population to support a seed orchard was used as a criterion.

61
Genetic gain Breeding population size =48, SPM, progeny size = 50, GMS selection, high heritability, after five generations 0 10 20 30 40 50 60 70 80 0246810121416 Gene diversity (status number) in the breeding population Seed orchard (production population) Breeding population based on Rosvall 1999 Conclusions: * Gene diversity in the breeding population makes it more able to support production populations Much unbalance No unbalance
 in the breeding population Seed orchard (production population) Breeding population based on Rosvall 1999 Conclusions: * Gene diversity in the breeding population makes it more able to support production populations Much unbalance No unbalance")
62
Unbalances in long term breeding Some advantage of unbalance is found, but so marginal and uncertain (Rosvall 1999) that it seemed doubtful applying unbalance in the Swedish long term breeding. Jag tror en moderat obalans i långsiktig förädling ger 3-7% bättre fröplantager det närmaste seklet jämfört med ”fundamentalistisk” balans.

63
Some reasons Rosvall (1999) found little advantage of unbalance The benefit of the breeding is measured as its ability of supporting seed orchards; Less sophisticated selection for advanced generation seed orchards than will be used in practice; Testing of the recruitment population (clonal or progeny), thus high heritability (Swedish pine breeding may turn to phenotypic selection next cycle, when unbalance may appear more favorable); Not exactly optimal unbalance, constraints in the simulator makes it hard to use optimum; Intensively selected breeding populations (Bulmer effect); Distinct generations, that will not be so!; Breeding population size is small, that makes conservation of diversity relatively important; Mainly a closed breeding population. I guess the advantage of unbalance is slightly greater in “real world”, and in particular in initiation!

64
Relative diversity 0.5 0 0 1 Gain Combined index=estimated BV (maximizes gain) Between family (exhausts diversity) Within family (preserves diversity) Gain at a given “diversity”. h 2 =0.25 and P=0.1 Modified From Lindgren and Wei 1993 Within family selection does not look efficient. Information from sibs was used for estimating breeding values (selection index). Infinite normal populations were assumed.
. Infinite normal populations were assumed..")
65
Distinct generations rolling synchronously will not work! Rolling front breeding is more operational, and must be unbalanced! Of the Swedish breeding populations, which reached F 1 in field, 75% are not synchronized. Thus take it as a fact of life that synchronization seldom works! Mates can be selected in several ways. Trees in field trials can probably only function as pollen parents, while grafts may first be available as seed parents. Some materials will be remeasured at higher age, some not, different materials develop differently; The management of rolling front will be unbalanced anyway, so that balance is simple is practically irrelevant even if mentally convenient.

66
Genetic value of component Contribution of component AB Equal resources More resources for improving larger components may result in better production populations! A B More resources for better Not only contributions but also resource allocation matters!

67
More attention on the better may improve efficiency If the predicted best contributions get more attention (e.g. larger test families) the best contributions benefit more from breeding. Seed orchards are drawn from the best part of the breeding population and benefits from such an unbalance.
 the best contributions benefit more from breeding. Seed orchards are drawn from the best part of the breeding population and benefits from such an unbalance..")
68
After Mullin et al 2005 Low Family breeding value High Family size Strong unbalance gives 20 % more gain Intermediate unbalance, 10% more gain Results interpreted from PhD thesis Lstiburek (2005). POPSIM simulations. Linear deployment of family sizes related to their breeding value combined with PAM and within family selection boosts ability to select for high gain seed orchards!

69
Why not stronger unbalance? It does not look like good breeding economy to spend lots of resources to produce a family (including their parents) and when make the family size very small for some families; Test environments and optimal test criteria for “optimizing” family size is different from there families are deployed, thus the advantage will be reduced; Allthough seed orchards benefit, the average of the breeding population will be reduced. The accepted conventional wisdom is the same family size, safer not to make too extreme changes, while experience and considerations accumulate successively stronger unbalance may be applied.
 and when make the family size very small for some families; Test environments and optimal test criteria for optimizing family size is different from there families are deployed, thus the advantage will be reduced; Allthough seed orchards benefit, the average of the breeding population will be reduced. The accepted conventional wisdom is the same family size, safer not to make too extreme changes, while experience and considerations accumulate successively stronger unbalance may be applied..")
70
Average selection intensity The selection gain by within family selection drops if the same total testing effort is unequally distributed among families, but the loss is marginal if the unbalance is moderate. Balanced Moderate unbalance Strong unbalance Sizei i i Large family51.16361.26791.485 Small family51.16341.02910 Average selection intensity 1.163 1.148 0.742 % of balanced 100 98.7 63.8

71
My conclusion from Lstiburek (2005): offspring should be larger from better parents even with balanced breeding, but only moderate unbalance is motivated, not strong. Note that Lstibureks results were obtained for a ”balanced” within family selection, but with ”unbalanced” allocation of resources. A more ”practical” algorithm could be written.

72
Population structuring Ph-thesis Rosvall (1999), Lstiburek (2005) To structure a breeding population in elite and main offers advantages, but more advantages of the same type can be achieved by proper management of a single population. Thus elite and main are not needed and should be replaced by algorithms with a less distinct border.

73
Stratified sublining From Ruotsalainen PhThesis (2001) Extends the PAM concept to several generations; A better alternative to “elite – main” is to use many strata.
 Extends the PAM concept to several generations; A better alternative to elite – main is to use many strata.")
74
Regeneration 60 (…100) Index breeding value HIGHEST LOWEST Stratified sublining for a breeding population in a region Breeding population F1 families Taken from Finnish breeding strategy (Haapanen 2004)
 Index breeding value HIGHEST LOWEST Stratified sublining for a breeding population in a region Breeding population F1 families Taken from Finnish breeding strategy (Haapanen 2004)")
75
Stratified sublines –stratification allows prioritising testing and breeding efforts on the sublines which are most likely contribute trees to seed orchards Complete control of inbreeding –enables enough unrelated selections to be deployed in seed orchards Flexible –sublines can be merged or entirely abandoned if desired Taken from Finnish breeding strategy (Haapanen 2004)
")
76
Parents are paired for PAM or allocated to stratified sublines based on certain optimal indices, but the optimal indices will be different in the offspring (different environment, different genetic parameters, different desires), thus the positive effect of PAM and stratified sublines probably is slightly overestimated. Slight overestimate of advantages

77
End

Liknande presentationer